
Les mycorhizes




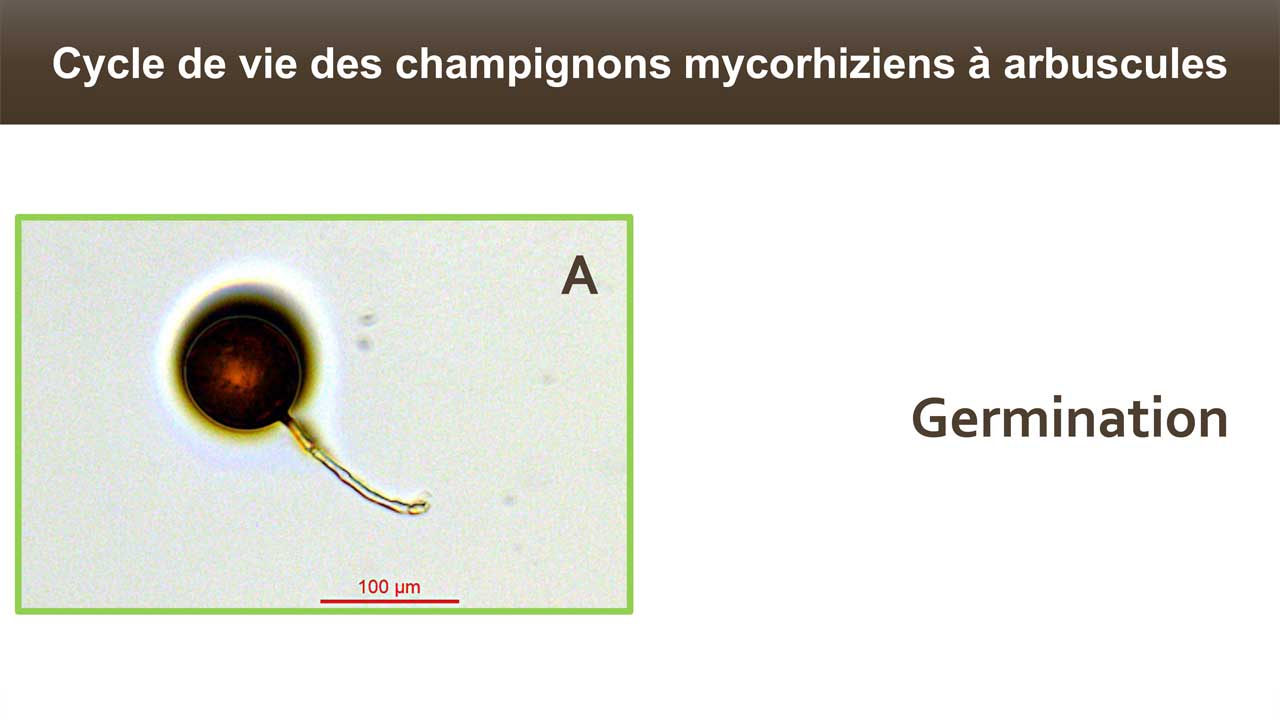


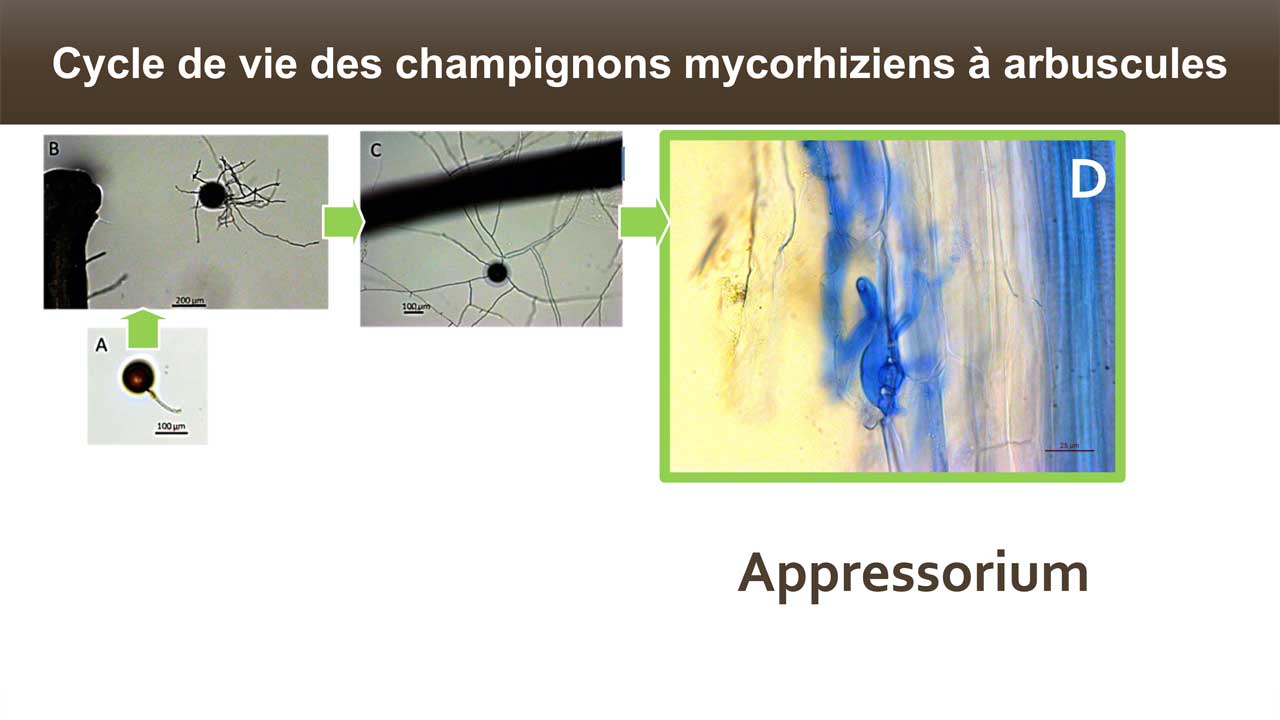





Le terme « Mycorhize » caractérise une relation intime et stable (symbiose mutualiste) entre des champignons et les racines des plantes. Ces symbioses génèrent un énorme réseau d’hyphes dans le sol, qui va s’associer à la communauté végétale et permettre l’apport d’eau, de nutriments aux plantes, améliorant ainsi leurs croissances et leurs résistances aux stress.
L’association mycorhizienne est l’association la plus importante dans les différents écosystèmes de notre planète en terme d’individus concernés et en terme d’impact sur les écosystèmes.
Il existe différents types de mycorhizes. Notre entreprise s’intéresse aux mycorhizes les plus anciennes et les plus répandues : les Mycorhizes à Arbuscules. En effet, ces mycorhizes colonisent 72% des plantes terrestres. Ces mycorhizes sont également les plus ancestrales puisqu’elles sont datées à plus de 450 millions d’années. Cette datation a été estimée grâce à la découverte de spores fossiles. Ces associations symbiotiques auraient joué un rôle prédominant dans l’évolution des plantes aquatiques en plantes terrestres (vascularisation).
La consommation en ressources primaires (minérales, eau, végétaux) ne cesse d’augmenter avec le développement technologique, économique et l’expansion industrielle et démographique.La croissance de la société et de la civilisation s’appuie principalement sur l’exploitation minière et l’exploitation agricole pour trouver les ressources indispensables à leur confort et à leurs besoins.
D’un côté l’exploitation minière détruit les écosystèmes présents sur les zones exploitées, ce qui conduit à la disparition de nombreuses populations animales, végétales et microbiennes, menaçant certaines espèces de disparition. Pour éviter cette perte de biodiversité, des programmes de restauration écologique doivent être mis en place.
D’un autre côté, l’agriculture moderne privilégie l’apport de phosphore et d’autres éléments essentiels sous leurs formes minérales, rapidement assimilables par les plantes ainsi que l’utilisation de pesticides aux dépens d’autres méthodes plus respectueuses de l’environnement.
Par conséquent certaines ressources naturelles s’épuisent devant une consommation grandissante des sociétés contemporaines et des problèmes sanitaires et environnementaux émergent liés aux méthodes utilisées par l’industrie minière et agricole. Des solutions doivent être apportées pour limiter l’utilisation de ces produits, tout en augmentant la production agricole qui selon les estimations devra augmenter d’environ 100% au cours du 21e siècle pour répondre aux demandes alimentaires de la population mondiale (Cirera et Masset 2010).
Par exemple, le phosphate est une ressource fossile utilisée massivement en agriculture et dans une moindre mesure pour la restauration écologique. Si le rythme d’extraction actuel se maintient, les réserves mondiales de phosphate seront épuisées d’ici 2100 (Cordell et al. 2009). Des solutions alternatives doivent être développées pour réduire cette consommation effrénée de phosphate et éviter une famine mondiale (FAO, 2014).
Une autre préoccupation environnementale majeure concerne les changements climatiques. En effet l’augmentation de la température globale terrestre cause un effondrement de la biodiversité, des extinctions d’espèces et des pertes d’individus composant les espèces survivantes (Thomas et al. 2004). Ce réchauffement climatique est causé par l’émission de gaz à effet de serre (Cox et al. 2000). L’agriculture intensive et l’exploitation minière génèrent des quantités importantes de gaz à effet de serre : dioxyde de carbone (CO2), méthane (CH4) et protoxyde de nitrate (N2O) (Soussana et al. 2007). L’apport d’engrais minéraux azotés (N) dans les systèmes agricoles est considéré comme la principale source d’émissions de N2O dans l’atmosphère (Mosier et al. 1998 ; Pattey et al. 2007). À l’échelle mondiale, les sources anthropiques de rejet de CO2, N2O et de CH4 dans l’atmosphère sont provoquées principalement par l’agriculture intensive et l’industrie extractive (Robertson et Grace, 2004). Ce phénomène risque de s’accentuer face à une augmentation de la population humaine qui selon les prévisions atteindra 10 milliards d’individus en 2100 (Lutz et al. 2001) et qui entraînera inévitablement l’expansion des zones agricoles et minières, selon les estimations plus d’un milliard d’hectares d’écosystèmes naturels (forêts) devraient disparaître d’ici 2050 (Tilman et al. 2001). Les forêts font partie des lieux de stockages du CO2 les plus importants de la planète (Lal et al. 2005). Cette augmentation d’émission de gaz à effet de serre est donc principalement causée par la déforestation (dégradation) des sols, liée à la nécessité d’augmenter l’espace agricole et minier pour subvenir aux besoins d’une population en pleine croissance démographique et par l’utilisation abusive d’intrants chimiques (engrais minéral azoté, phosphaté et pesticides) pour l’agriculture intensive et dans une moindre mesure pour la restauration écologique qui dégradent la qualité des sols limitant ainsi le stockage du CO2 dans les terres et entraînant un relargage important de N2O dans l’atmosphère (Tilman et al. 2001 ; Pattey et al. 2007).
Notre civilisation en pleine croissance démographique entraîne également une demande croissante en matière première et en énergie. Pour parvenir à répondre à cette demande croissante, une augmentation constante du rythme d’extraction par l’industrie minière des ressources minérales issues des gîtes métallifères est nécessaire (Harrison & Rajakaruna 2011). La plupart des gîtes métallifères se trouve dans les manteaux d’altération latéritique de la ceinture tropicale. C’est le cas notamment en Nouvelle-Calédonie où les sols ultramafiques issus de ces manteaux d’altération ont des teneurs élevées en métaux et des carences en éléments essentiels pour les plantes. Ces conditions édaphiques extrêmes engendrent une biodiversité exceptionnelle (Barthlott et al. 1996 ; Brooks 1987 ; Proctor 2003), dont une partie est en danger d’extinction en raison de pressions anthropiques diverses (Myers et al. 2000). L’industrie minière, notamment, détruit ces écosystèmes, induisant une perte de biodiversité, la pollution des rivières et des océans et l’émission de gaz à effet de serre.
De plus, les organismes vivants dans des conditions environnementales extrêmes comme celles des milieux ultramafiques ont des traits uniques et sont donc extrêmement intéressants pour la découverte d’organismes ayant des intérêts biotechnologiques. Il est impératif de mettre en place des mesures de préservation et restauration écologique pour protéger et conserver la biodiversité de ces écosystèmes, afin de limiter entre autres, les pollutions engendrées par l’érosion et le lessivage des sols. Pour restaurer ces écosystèmes dégradés, les processus écologiques initiaux doivent être étudiés afin de pouvoir reconstituer une interface entre les sols et les plantes proche de l’origine du point de vue de son fonctionnement (Dobson 2008).
Une des solutions les plus prometteuses pour augmenter la productivité des plantes tout en diminuant l’utilisation d’intrants chimiques, et donc limiter les émissions des gaz à effet de serre dans les écosystèmes agricoles ou à restaurer, est l’utilisation des micro-organismes biostimulants des plantes tels que les champignons mycorhiziens à arbuscules (CMA) (Berruti et al. 2016). En effet, ces champignons ont prouvé leur efficacité dans des contextes agronomiques et écologiques en limitant les apports d’intrants chimiques (pesticides, N, P, K) et en conférant aux plantes une meilleure nutrition minérale et une tolérance aux stress biotiques et abiotiques du milieu. Les CMA jouent également un rôle vital dans le cycle global du carbone, car ils peuvent utiliser une grande partie du carbone fixé par les plantes, produire des composés organiques à cycle lent (glomaline) et protéger la matière organique contre les attaques microbiennes en favorisant l’agrégation des sols (Giri et Saxena 2017). Parmi les associations plantes-champignons, la plus fréquente est la symbiose mycorhizienne à arbuscules : la majorité des espèces végétales sont associées, au niveau de leur racine, avec des champignons du groupe des Gloméromycètes. Cette symbiose est retrouvée sur tous les continents des îles subarctiques à la Péninsule antarctique et dans tous les écosystèmes terrestres (Newsham et al. 2009; Smith et Read 2008). La quasi-universalité de cette symbiose tient au bénéfice nutritionnel que les plantes en retirent depuis plus de 400 millions d’années. En effet, les champignons mycorhiziens à arbuscules (CMA) auraient permis de promouvoir l’apparition de racines chez les plantes (Fortin et al. 2015), ce qui aurait permis la conquête des milieux terrestres par les plantes permettant ainsi de transformer les milieux terrestres en milieux fertiles, c’est-à-dire en sol vivant. On doit à ces tous premiers sols l’établissement graduel des végétaux et, conséquemment, l’existence du monde vivant, animal aussi bien que végétal, sur la surface terrestre (Fortin et al. 2015). De même que les animaux ne peuvent correctement digérer leur bol alimentaire qu’en étant associé à une flore microbienne présente dans leur tube digestif, les plantes, à part quelques exceptions, ne peuvent efficacement retirer les sels minéraux du sol qu’en étant associées à des champignons mycorhiziens.
La symbiose mycorhizienne à arbuscule est invisible à l’oeil nu. Aussi, nous ne nous doutons pas de l’abondance de cette symbiose dans notre environnement : un cm3 de sol de prairie renferme plusieurs dizaines de mètres d’hyphes (Ji et al. 2013). Le champignon forme un réseau mycélien à l’extérieur des racines, explorant le sol et puisant eau et sels minéraux, particulièrement le phosphore et dans une moindre mesure l’azote. Ces éléments, présents dans le sol mais sous forme peu mobile (P) ou peu abondante (N), sont transférés à la plante hôte en contrepartie de glucose (issu de la photosynthèse) et de lipide (Keymer et al. 2017). Près de 20% du carbone photosynthétique est transféré au champignon symbiotique (Jakobsen & Rosendahl 1990). Dans la nature 90% du phosphore et 60% de l’azote présents dans les plantes sont issus de l’activité de ces champignons (Smith et Read 2008). En étendant la capacité d’exploration des racines dans le sol, le réseau mycélien améliore la nutrition minérale et hydrique des plantes.
Pour toutes ces raisons, l’homme doit s’il veut pérenniser la vie sur Terre, remplacer le plus vite possible les produits néfastes pour l’environnement (engrais chimiques et pesticides) par des solutions naturelles tel que les CMA pour la culture des plantes afin de conserver une productivité suffisante et de régénérer les sols dégradés.
Bibliographie :
Barthlott W, Lauer W, Placke A (1996) Global distribution of species diversity in vascular plants: towards a world map of phytodiversity. Erdkunde 50:317-326.
Berruti A, Lumini E, Balestrini R, Bianciotto V (2016) Arbuscular Mycorrhizal Fungi as Natural Biofertilizers: Let’s Benefit from Past Successes. Front Microbiol 6:. https://doi.org/10.3389/fmicb.2015.01559.
Brooks RR (1987) Serpentine and its vegetation: a multidisciplinary approach. Dioscorides Press, Portland 454 p.
Cirera X, Masset E (2010) Income distribution trends and future food demand. Philosophical Transactions of the Royal Society B: Biological Sciences 365:2821–2834. https://doi.org/10.1098/rstb.2010.0164
Cordell D, Drangert J-O, White S (2009) The story of phosphorus: Global food security and food for thought. Global Environmental Change 19:292–305. doi:10.1016/j.gloenvcha.2008.10.009
Dobson C (2008) Ecological field studies: through field studies, students learn scientific process and ecological content. The Science Teacher 2/01/2008.
Fortin JA, Plenchette C, Piché Y (2015) L’essor de la nouvelle révolution verte. Quae.com. http://www.quae.com/fr/r4798-les-mycorhizes.html.
Giri B, Saxena B (2017) Response of Arbuscular Mycorrhizal Fungi to Global Climate Change and Their Role in Terrestrial Ecosystem C and N Cycling. In: Varma A, Prasad R, Tuteja N (eds) Mycorrhiza – Function, Diversity, State of the Art. Springer International Publishing, Cham, pp 305–327
Harrison S, Rajakaruna N (2011) Serpentine: The Evolution and Ecology of a Model System. Berkeley, CA: University of California Press, Orlando, FL, USA.
Jakobsen I, Rosendahl L (1990) Carbon flow into soil and external hyphae from roots of mycorrhizal cucumber plants. New Phytol 115: 77–83.
Ji B, Gehring CA, Wilson GWT, Miller RM, et al (2013) Patterns of diversity and adaptation in Glomeromycota from three prairie grasslands. Mol Ecol 22: 2573–2587.
Keymer A, Pimprikar P, Wewer V, et al (2017) Lipid transfer from plants to arbuscular mycorrhiza fungi. eLife Sciences 6:e29107. doi: 10.7554/eLife.29107
Lutz W, Sanderson W, Scherbov S (2001) The end of world population growth. Nature 412:543–545. https://doi.org/10.1038/35087589
Lal R (2005) Forest soils and carbon sequestration. Forest Ecology and Management 220:242–258. https://doi.org/10.1016/j.foreco.2005.08.015
Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853-858.
Newsham K-K, Upson R, Read D-J. 2009. Mycorrhizas and dark septate root endophytes in polar regions. Fungal Ecol 2, 10-20.
Mosier A, Kroeze C, Nevison C, et al (1998) Closing the global N2O budget: nitrous oxide emissions through the agricultural nitrogen cycle. Nutrient Cycling in Agroecosystems 52:225–248. https://doi.org/10.1023/A:1009740530221
Pattey E, Edwards GC, Desjardins RL, et al (2007) Tools for quantifying N2O emissions from agroecosystems. Agricultural and Forest Meteorology 142:103–119. https://doi.org/10.1016/j.agrformet.2006.05.013
Proctor J (2003) Vegetation and soil and plant chemistry on ultramafic rocks in the tropical Far East. Perspectives in Plant Ecology, Evolution and Systematics 6:105-124
Robertson GP, Grace PR (2004) Greenhouse Gas Fluxes in Tropical and Temperate Agriculture: The Need for a Full-Cost Accounting of Global Warming Potentials. In: Wassmann R, Vlek PLG (eds) Tropical Agriculture in Transition — Opportunities for Mitigating Greenhouse Gas Emissions? Springer Netherlands, Dordrecht, pp 51–63
Smith SE, Read DJ. 2008. Mycorrhizal Symbiosis, Third Edition. Academic Press.
Soussana JF, Allard V, Pilegaard K, et al (2007) Full accounting of the greenhouse gas (CO2, N2O, CH4) budget of nine European grassland sites. Agriculture, Ecosystems & Environment 121:121–134. https://doi.org/10.1016/j.agee.2006.12.022
Thomas CD, Cameron A, Green RE, et al (2004) Extinction risk from climate change. Nature 427:145–148. https://doi.org/10.1038/nature02121
Tilman D, Cassman KG, Matson PA, et al (2002) Agricultural sustainability and intensive production practices. Nature 418:671–677. https://doi.org/10.1038/nature01014